BIOLOGÍA Y GEOLOGÍA
LA CÉLULA: MORFOLOGÍA CELULAR
1. BIOMEMBRANAS
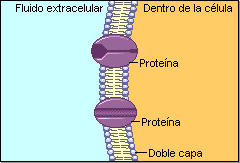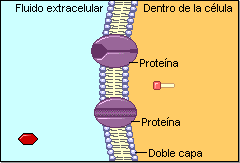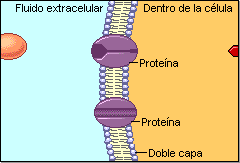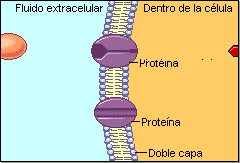
Las biomembranas son láminas fluidas, encargadas de separar el interior celular de su medio externo, así como de delimitar los diferentes orgánulos citoplasmáticos. Son barreras de permeabilidad selectiva, que controlan las entradas y salidas de sustancias y mantienen unas condiciones estables en el interior celular. Las membranas tienen un grosor de unos 6-10 nm, por lo que no pueden verse al microscopio óptico. Con el electrónico tienen el aspecto de una doble lámina oscura separada por una banda clara.1.1 ESTRUCTURA DE LAS BIOMEMBRANAS
El modelo de membrana aceptado actualmente fue propuesto en 1972 por Jonathan Singer y Garth Nicholson y se denomina de mosaico fluido. Según este modelo, las membranas están constituidas principalmente por una bicapa lipídica, componente mayoritario, y proteínas distribuidas en esa bicapa. Las proteínas son las que dan especificidad a las funciones de las membranas. La proporción entre lípidos y proteínas es variable. En la plasmática suele ser al 50%. En mitocondrias hay un 80% de proteínas. En las vainas de mielina de los axones nerviosos las proteínas sólo representan un 18%.1.2.LA BICAPA LIPÍDICA: ESTRUCTURA Y PROPIEDADES
La bicapa lipídica está formada por fosfolípidos (mayoritarios), glucolípidos y colesterol, en proporciones variables. El carácter anfipático de todos estos lípidos le proporciona a las membranas una serie de características: • Autoensamblaje: en medio acuoso se forman bicapas de forma espontánea, ya que las regiones hidrófilas quedan en contacto con el agua y las hidrófobas en el interior. • Autosellado: las bicapas tienden a cerrarse sobre sí mismas, formando vesículas esféricas. • Fluidez: las bicapas son fluidas, permitiendo el movimiento de las proteínas y de los lípidos dentro de una misma capa. Sin embargo, el paso de un fosfolípido (o una proteína) de una monocapa a la otra (movimiento flip-flop) no se da, por lo que ambas monocapas son distintas: asimetría lipídica. La fluidez de la membrana depende, entre otros factores, de la cantidad de colesterol que contenga. A mayor cantidad mayor rigidez y resistencia. Por ello, las membranas pueden alterar la composición lipídica (más colesterol, saturaciones y longitud de los ácidos grasos) para compensar cambios de temperatura. • Impermeabilidad: debido a su carácter hidrofóbico, las membranas son bastante impermeables a iones y moléculas hidrosolubles de cierto tamaño. Así, se puede impedir la entrada y salida de sustancias hidrosolubles.1.3.PROTEÍNAS DE MEMBRANA
Las proteínas confieren las propiedades funcionales a cada membrana. Según su posición en la membrana, las proteínas se clasifican en integrales y periféricas. • Proteínas integrales: unidas fuertemente a los lípidos. Las proteínas de transmembrana atraviesan la membrana una vez (de paso único) o varias veces (multipaso). Otras están por fuera, pero unidas covalentemente a los lípidos. • Proteínas periféricas: a un lado u otro de la membrana. Presentan uniones no covalentes con las cabezas polares de los lípidos o con proteínas integrales. Muchas proteínas son glucoproteínas, ya que llevan oligosacáridos. Se forman en la luz del retículo endoplasmático o del aparato de Golgi, por lo que siempre aparecen en la cara no citosólica de la membrana. Al igual que los lípidos, las proteínas se desplazan por las membranas, pero no pueden invertir su posición: asimetría proteica.2. LA MEMBRANA PLASMÁTICA
La membrana plasmática limita y comunica el interior celular del exterior. Presenta una estructura típica en mosaico fluido. En su cara externa muestra una cubierta fibrosa que no aparece en otras membranas. Es el glucocálix, formado por los oligosacáridos de glucolípidos y glucoproteínas. El glucocálix protege de daños físicos y químicos, filtra sustancias y es un medio de comunicación, reconocimiento y adhesión celular. La membrana plasmática controla la entrada y salida de sustancias (permeabilidad selectiva), interviene en las uniones celulares que permiten la formación de tejidos y recibe y transmite señales.2.1. PERMEABILIDAD SELECTIVA
La permeabilidad selectiva permite a la célula controlar las entradas y salidas y, por tanto, su medio interno. El carácter lipídico de la membrana la hace impermeable a numerosas sustancias, por lo que posee un complejo sistema de transporte específico realizado por proteínas de membrana. Este sistema permite el paso de sustancias de diferentes tamaños.A) Transporte de moléculas pequeñas
Puede ser pasivo (sin gasto de energía) o activo, que precisa energía. A.1) Transporte pasivo o difusión Las moléculas se mueven espontáneamente a favor de gradiente (de mayor a menor). Existen 2 tipos: a) Difusión simple: a través de la bicapa lipídica. La realizan moléculas no polares: gases (O 2 , CO 2 ), hormonas liposolubles (esteroideas, tiroideas). Las moléculas polares muy pequeñas también pueden realizarla: agua, etanol, glicerol, urea. b) Difusión facilitada. Requiere de proteínas transportadoras (permeasas) o de proteínas canal. Se utiliza para iones y grandes moléculas polares. b.1) Proteínas transportadoras o permeasas. Son proteínas transmembrana que se unen específicamente a ciertas sustancias. Esta unión altera la estructura proteica y permite el transporte. Luego, la proteína recupera su estructura original. Cada molécula transportada (azúcares, aminoácidos, metabolitos, etc) tiene su propio transportador. b.2) Proteínas canal o canales iónicos. Proteínas transmembrana que forma poros acuosos para el paso de Na + , Ca 2+ , Cl - . Los canales iónicos permanecen cerrados hasta que una señal externa los abre. En los canales iónicos dependientes de ligando la señal es química (hormonas, neurotransmisores,…). En los canales iónicos dependientes de voltaje la señal es eléctrica (potencial de membrana). A.2) Transporte activo Las moléculas atraviesan las membranas en contra de gradiente, para lo que precisan energía, habitualmente en forma de ATP. Lo realizan proteínas transportadoras denominadas bombas. Una de las más conocidas es la bomba de Na + /K + . La energía de hidrólisis del ATP se utiliza para expulsar 3 iones de Na + e introducir 2 de K + , ambos contra gradiente. Así se puede controlar la ósmosis o producir potenciales de membrana.B) Transporte de macromoléculas y partículas
Este tipo de transporte incluye la entrada (endocitosis) y la salida (exocitosis) de sustancias mediante la formación de vesículas, sacos membranosos que se desplazan por el citoplasma. B.1) Endocitosis Las sustancias se engloban en invaginaciones de la membrana que se cierran formando vesículas intracelulares. Los procesos mejor conocidos son la fagocitosis y la endocitosis dependiente de clatrina. • Fagocitosis: se ingieren partículas muy grandes (bacterias, células, orgánulos,…). Contra invasores o para eliminar células propias. La membrana forma prolongaciones llamadas pseudópodos que rodean a la partícula a fagocitar, formando una gran vesícula o fagosoma. Los materiales fagocitados serán digeridos por los lisosomas. Para líquidos se da pinocitosis, con invaginaciones y sin pseudópodos. • Endocitosis dependiente de clatrina: la entrada es selectiva. Las vesículas que englobarán a las partículas se forman en regiones especiales de la membrana, fosas cubiertas, depresiones que tienen su cara citosólica cubierta por la proteína clatrina. El proceso es similar, formándose vesículas endocíticas que se convierten en endosomas que acabarán en los lisosomas. Un ejemplo típico es la captación de colesterol. Los dos tipos de endocitosis están mediados por proteínas de la membrana, los receptores de endocitosis, que reconocen las partículas o macromoléculas específicas a ingerir. B.2) Exocitosis Proceso inverso a la endocitosis. Permite a las células secretar sustancias para renovar la membrana plasmática y la matriz extracelular, así como la secreción de hormonas neurotransmisores, enzimas digestivos, etc, en células especializadas. Los productos a secretar se fabrican en el retículo endoplasmático y luego pasan al aparato de Golgi. Allí pueden modificarse y son englobados en vesículas secretoras que se dirigirán a la membrana plasmática para su expulsión.3. EL CITOPLASMA
El citoplasma es el contenido celular entre la membrana y el núcleo (en células eucariotas). Contiene una porción acuosa (citosol), con numerosos filamentos proteicos (citoesqueleto) y gran variedad de estructuras y orgánulos.4. EL CITOSOL
El citosol, citoplasma fundamental o hialoplasma es la porción líquida del citoplasma. Contiene básicamente agua en la que están inmersos los orgánulos membranosos, los ribosomas, enzimas, inclusiones y el citoesqueleto. En el citoplasma tiene lugar la síntesis, plegamiento y degradación de numerosas proteínas, así como la mayoría de las reacciones metabólicas (glucólisis, gluconeogénesis, glucogenolisis, etc).5. INCLUSIONES CITOPLASMÁTICAS
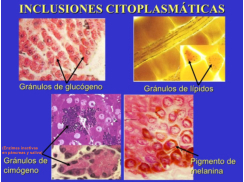
Son materiales almacenados en el citoplasma y no rodeados de membranas. Destacan el glucógeno y las grasas (reservas de energía). • Glucógeno: polímero ramificado de glucosas. Al microscopio electrónico aparece como gránulos dispersos, sobre todo en células del hígado y músculo. • Grasas: fuente de energía más importante. Proporciona el doble de energía por gramo que los glúcidos y, además, ocupa menor espacio. La mayor parte de las grasas se almacenan en el citoplasma del tejido adiposo, como una gran gota de triglicéridos.6. LOS RIBOSOMAS
Son orgánulos celulares no membranosos. Máquinas moleculares encargadas de sintetizar proteínas. Están compuestos de ARNr y más de 50 proteínas. Los ribosomas se denominan por su coeficiente de sedimentación, medida en unidades Svedberg (1S=10 -13 segundos). Los procariotas son 70 S y los eucariotas, 80 S. Ambos tienen dos subunidades, una grande y otra pequeña. En eucariotas los ribosomas pueden hallarse en: la cara citosólica de la membrana nuclear externa y de la membrana del retículo endoplasmático rugoso, libres en el citoplasma y en el interior de mitocondrias y cloroplastos. Para sintetizar proteínas, los ribosomas se unen al ARNm en grupos espirales llamados polirribosomas o polisomas. Los tripletes del ARNm son leídos y traducidos a proteínas con los aa que transportan al ribosoma los diferentes ARNt.7. EL CITOESQUELETO
El citoesqueleto, estructura que se creía exclusiva de los eucariotas pero se ha demostrado presente en procariotas, está formado por una red de varios tipos de filamentos proteicos, se extiende por todo el citoplasma y se ancla a la membrana. Existen tres tipos de filamentos en el citoesqueleto: microtúbulos, microfilamentos (o filamentos de actina) y filamentos intermedios. Todos ellos interaccionan entre sí, con los orgánulos celulares y con la membrana mediante proteínas accesorias. El citoesqueleto es una estructura cambiante, pues se modifica según las necesidades de la célula y en la división.7.1. COMPONENTES DEL CITOESQUELETO
Microtúbulos y microfilamentos: son estructuras polares, es decir, que sus dos extremos son diferentes. Uno de los extremos crece (o decrece) a gran velocidad, añadiendo monómeros de actina (microfilamentos) o de tubulina (microtúbulos). El otro lo hace de forma lenta. El extremo rápido se denomina extremo más (+) y el lento extremo menos (-). Son estructuras lábiles, es decir, pueden crecer o decrecer añadiendo o eliminando monómeros proteicos en sus extremos. La estabilidad depende de proteínas asociadas. Microtúbulos y microfilamentos pueden formar parte de estructuras dinámicas (anillo contráctil de citocinesis, pseudópodos, huso mitótico) y estables (sarcómeros, microvellosidades, cilios, flagelos). Los microfilamentos se extienden por todo el citoplasma, pero abundan bajo la membrana plasmática: córtex celular. Participan en el mantenimiento de la forma celular, los pseudópodos, el anillo contráctil de la citocinesis y la contracción muscular. Los microtúbulos irradian desde el centrosoma hasta la periferia nuclear. Participan en la forma celular, cilios y flagelos, movimiento de orgánulos y huso acromático. Filamentos intermedios: son estructuras estables, no polares. Su composición es variada, por lo que reciben diferentes nombres según las células en que se hallan: queratina (tejido epitelial), vimentina (conjuntivo y muscular), neurofilamentos (nervioso).7.2. CENTROSOMA
Es el centro organizador de microtúbulos (COM), pues los fabrica, coloca y orienta. El centrosoma está formado por una matriz amorfa con cientos de anillos proteicos de un tipo especial de tubulina que inicia la nucleación de los microtúbulos. Éstos se unen a los anillos por su extremo (-) y crecen por su extremo (+), irradiando desde el centrosoma.7.3. ESTRUCTURAS FORMADAS POR MICROTÚBULOS
7.3.1. CENTRIOLOS Son dos estructuras cilíndricas (0,25 de anchura y de 0,15 a 0,5 de altura), perpendiculares entre sí y embebidas en el centrosoma de células animales. El conjunto se denomina diplosoma. Cada centriolo consta de 9 tripletes de microtúbulos, llamados A, B y C, de los que sólo el A está completo (Estructura 9+0). Además, contienen numerosas proteínas accesorias que conectan los tripletes entre sí y con el centro del centriolo. Centriolos y centrosoma se duplican en cada ciclo celular al mismo tiempo que el ADN, antes de la mitosis. Al comenzar ésta, cada diplosoma se dirige a uno de los polos de la célula arrastrado por el huso mitótico. 7.3.2. CILIOS Y FLAGELOS Son prolongaciones de la membrana plasmática formadas por microtúbulos y proteínas asociadas. Son responsables del movimiento de ciertas células. Si las células son libres, el movimiento de cilios y flagelos desplaza a la propia célula. En células fijas, se producen corrientes en el líquido que las baña. Cilios y flagelos tienen una estructura similar. Sin embargo, se mueven de forma diferente y, además, los cilios son cortos, numerosos y delgados, mientras que los flagelos son pocos, largos y gruesos. ESTRUCTURA DE CILIOS Y FLAGELOS Cilios y flagelos constan de una parte exterior al cuerpo celular cubierta por la membrana plasmática (el axonema) y otra porción interna, bajo la membrana plasmática (el cuerpo basal). El cuerpo basal tiene la misma estructura que el centriolo (9+0) y es el que organiza el cilio y del que crece el axonema. El axonema tiene una estructura 9 + 2: 9 pares de microtúbulos periféricos y un par central. Los dos microtúbulos centrales son completos, pero los pares periféricos contienen uno completo (A) y otro incompleto (B). El axonema también contiene muchas proteínas con función estructural o motora. Así, la nexina une los pares de microtúbulos externos entre sí. La dineína, en forma de dos brazos que salen del microtúbulo A, es una proteína motora, responsable de los movimientos de los cilios y flagelos con la energía que proporciona el ATP. MECANISMO DEL MOVIMIENTO DE CILIOS Y FLAGELOS El movimiento de cilios y flagelos se debe al desplazamiento de unos dobletes externos de microtúbulos sobre otros. En presencia de ATP, la dineína que sale del microtúbulo A de un doblete se desliza a lo largo del túbulo B del doblete contiguo. Esto provoca la flexión del cilio o flagelo y su movimiento de batido.




















Para ir a donde no se sabe hay que ir por donde no se sabe.” San Juan de la Cruz
“It must be a strange world not being a scientist, going through life not knowing--or
maybe not caring about where the air came from, where the stars at night came from or
how far they are from us. I WANT TO KNOW” Michio Kaku
“Nullius in verba” Robert Boyle, Christopher Wren y Robert Hooke










Vistas frontal y posterior de un modelo 3D de la región de Golgi en una célula de mamífero secretora de insulina. Se reconstruyeron, mediante tomografía EM
de doble eje, tres secciones seriales de 400 nm de grosor cortadas de una célula cogelada a alta presión, congelada con un sustituto y envueltas en plástico
HIT-T15. El software IMOD se usó para modelar todos los objetos visibles dentro del volumen reconstruido resultante (3,1 x 3,2 x 1,2 um
3
). El complejo de
Golgi con siete cisternas (C1-C7) está en el centro. La codificación de color es la siguiente: C1, azul claro; C2, rosa; C3, rojo cereza; C4, verde; C5, azul
oscuro; C6, oro; C7, rojo brillante. El Golgi se muestra en el contexto de todos los orgánulos, vesículas, ribosomas y microtúbulos circundantes: retículo
endoplásmico (ER), amarillo; ribosomas unidos a la membrana, azul; ribosomas libres, naranja; microtúbulos, verde brillante; vesículas de núcleo denso, azul
brillante; vesículas clatrina-negativas, blancas; compartimientos y vesículas clatrina-positivas, rojo brillante; compartimientos y vesículas negativas a clatrina,
púrpura; mitocondrias, verde oscuro. Bar, 500 nm
Image courtesy of Dr. Brad Marsh, Institute for Molecular Bioscience, The University of Queensland, Australia. Originally published in the Inaugural Article: Organellar
relationships in the Golgi region of the pancreatic beta cell line, HIT-T15, visualized by high resolution electron tomography. Proceedings of the National Academy of
Sciences USA (February 27, 2001)—volume 98, no. 5, pp 2399-2406. © 2001 National Academy of Sciences, U.S.A.










Animación de la bomba de NA/K. Iones de Na
+
en
amarillo; K
+
en verde; ATP en rosa.














Ejemplos de las tres vías de exocitosis. La vía
secretora constitutiva (flecha naranja) entrega
directamente lípidos y proteínas de membrana fresca
a la membrana celular. La vía regulada (flechas
verdes) se usa para carga específica como hormonas
y neurotransmisores. Las vesículas secretoras
circulan y se acumulan en la membrana, donde
esperan una señal para su liberación. La vía
lisosómica transporta directamente la carga a la
membrana (flecha roja), o digiere y expulsa los
desechos de la célula (flechas rosadas).








LA CÉLULA: MORFOLOGÍA CELULAR



Cómo se forman los pseudópodos



![[x]](bg2bto_htm_files/close.png "Close")